/ Главная / Разные статьи / Физиология / Вызванные потенциалы мозга
Вызванные потенциалы зрительной коры мозга при разных условиях световой стимуляции
A. E.Tambiev, P. N. Ermakov and Yu.V. Shcherbatykh. Evoked potentials in the rat visual cortex in different conditions of light stimulation. // Sechenov Physiological Journal of the USSR LXV, № 5, 1979, Р. 656-660.
А. Э. Тамбиев, П. Н. Ермаков, Ю. В. Щербатых. Вызванные потенциалы в зрительной коре мозга крысы при разных условиях световой стимуляции// Физиологический журнал СССР им. И. М. Сеченова. 1979. т.65, №5, С.656-660.
Аннотация: Изучались фокальные потенциалы, возникающие в зрительной коре мозга крысы при локальной световой стимуляции. Исследована динамика латентных периодов, амплитуды и длительности ответов в различных слоях коры, при увеличении диаметра стимулов и при смещении стимулов от области максимального ответа. На основании полученных данных представлена пространственная реконструкция распределения возбуждения в поле 17 зрительной коры на предъявление локальных стимулов.
Ключевые слова: фокальный потенциал, поверхностно-позитивное колебание, глубинно-негативное колебание, фокус возбуждения.
Согласно концепции об ансамблевой организации анализаторных структур мозга [3], основная роль в представлении и переработке информации принадлежит объединениям нервных клеток. Поэтому исследования распределения фокальных потенциалов, в известной степени отражающих суммарные процессы возбуждения и торможения [8], способствуют лучшему пониманию внутрикорковых механизмов переработки зрительной информации.
МЕТОДИКА
Опыты проведены на белых крысах (вес 100—150 г), обездвиженных внутримышечным введением d-тубокурарина (0.4 мг/кг). Фокальные потенциалы в первичной проекционной зрительной области коры, соответствующей центральной части сетчатки [8], регистрировались стеклянными микроэлектродами (электролит 3 мол. раствор КС1, сопротивление 0.8—2 мОм). Микроэлектроды вводились строго перпендикулярно по отношению к поверхности мозга через трепанационное отверстие в своде черепа. Глубина погружения определялась по шкале микрометра ММ-1, установленного на манипуляторе. Момент контакта микроэлектрода с поверхностью коры мозга отмечался по исчезновению наводки на экране осциллографа и визуально, под контролем микроскопа МБС-2. Усиление, наблюдение и регистрация фокальных потенциалов и импульсной активности нейронов осуществлялись на электрофизиологической установке УЭФПТ-5.
Сферический экран (фоновая освещенность около 10 Лк), на который проецировались световые стимулы, находился на расстоянии 30 см от глаза животного. Раздражения в виде вспыхивающих пятен различного диаметра (освещенность на единицу поверхности 70 Лк) подавались от стимулятора ФОС-2. Фокус возбуждения в коре определялся по минимальному латентному периоду фокального потенциала (при помещении тестирующего пятна в определенную точку экрана). Ипсилатеральный глаз закрывался, контралатеральный покрывался тонким слоем силиконовой жидкости для предохранения от подсыхания [9].
Во время эксперимента температура тела животного поддерживалась на постоянном уровне — 37° с помощью электроподогрева. На протяжении опыта проводился контроль функционального состояния животного по ЭКОГ и ЭКГ.
Фокальные потенциалы записывались на фотопленку с экрана осциллографа. Запуск ждущей развертки осуществлялся синхронно с подачей раздражителя.
На основании измерений латентных периодов, амплитуды и длительности глубинно-негативного колебания фокальных потенциалов строились эмпирические линии регрессии. Для узловых точек определялись средние ошибки Sx и доверительные границы х—tgs =g у. < x+tgj [6], уровень значимости р=0.05. Представлены данные по 32 микроэлектродным проходкам.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Распределение вызванных потенциалов от поверхности коры к белому веществу мозга.
От поверхности коры до глубины 200—300 мкм фокальный потенциал начинался поверхностно-позитивным колебанием. Средний латентный период данной фазы ответа (диаметр стимулов во всех случаях 5°) составил 30+2.5 мс, амплитуда варьировала от 200 до 400 мкВ, а длительность — от 15 до 30 мс. По мере погружения электрода в глубь коры амплитуда и длительность положительной фазы уменьшились и на глубине около 300 мкм наступала инверсия знака потенциала.
Амплитуда глубинно-негативной фазы потенциала достигала наибольших значений в нижней половине 3-го, 4-м и верхней половине 5-го слоя. Как следует из работы Н. И. Артюхиной [1] и некоторых других данных [10], разветвления специфических афферентов в зрительной коре мозга крыс и мышей приходятся именно на эти слои. В нижней части 5-го и 6-м слое (начиная с глубины 1200—1300 мкм) фокальные потенциалы уже практически отсутствовали. А. Я. Супин связывает это с более резким выявлением области «источников» и «стоков» синаптического тока [6]. Размер возникающего в коре диполя при локальной стимуляции существенно меньше, чем при диффузном освещении. Об этом свидетельствует также и распределение латентных периодов вызванных потенциалов.
Минимальные латентные периоды фокальных потенциалов регистрировались на глубине 600—800 мкм (4-й слой), где они составили в среднем 26+1.3 мс. В верхних слоях коры и в 5-м слое латентные периоды увеличились соответственно на 5—6 и на 2—3 мс в сравнении с минимальными значениями.
Амплитуда фокальных потенциалов была наибольшей в 4-м слое — 400+50 мкВ. В верхних и нижних слоях коры амплитуда была ниже — 250—300 мкВ. На глубине 1200—1300 мкм, как уже указывалось, амплитуда потенциалов резко падала до уровня фоновых колебаний.
Длительность фокальных потенциалов оказалась наиболее вариабельной характеристикой ответа. Однако усредненные данные по 16 микроэлектродным проходкам показали достоверное возрастание длительности фокального потенциала при погружении электрода от 300 до 900 мкм и его укорочение при дальнейшем погружении до 1200 мкм. Отметим несовпадение зоны максимальной амплитуды и максимальной длительности. По-видимому, это следствие синхронности афферентного залпа в воспринимающих слоях и более выраженной десинхронизации возбуждения на нейронах нижних слоев. Описанные зависимости представлены на рис. 1.
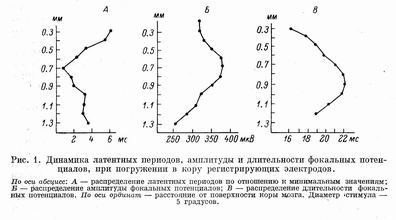
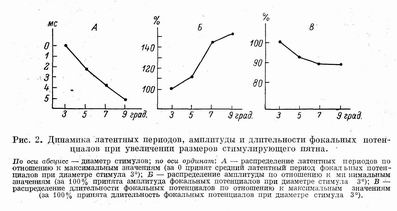
Распределение вызванных потенциалов при увеличении диаметра стимулов. Сопоставление ответов на локальные стимулы разного диаметра показало, что при увеличении светового пятна фокальные потенциалы закономерно увеличиваются. В диапазоне предъявляемых стимулов (от 3 до 9°) латентные периоды уменьшались в среднем на 5 мс, «скорость» укорочения при этом замедлялась по мере прироста диаметра светового пятна (от 3 до 5° — 2.2 мс, от 5 до 7° — 1.7 мс, от 7 до 9° — 1.3 мс).
Амплитуда фокальных потенциалов возрастала при увеличении диаметра стимулов. При увеличении размеров пятна от 3 до 9° прирост составил около 50%, но зависимость не была линейной. Максимум роста амплитуды потенциалов наблюдался при увеличении диаметра стимулов от 5 до 7°. Длительность фокальных потенциалов несколько уменьшается при увеличении диаметра светового пятна от 3 до 5°, а при дальнейшем увеличении стимула остается практически неизменной.
Уменьшение латентных периодов и увеличение амплитуды фокальных потенциалов при увеличении диаметра стимулов связаны с пространственной суммацией возбуждения на всех этапах переработки зрительной информации, благодаря чему растет синхронность и сила срабатывания нервных элементов [7]. Динамика длительности фокальных потенциалов, по-видимому, является результатом двух взаимосвязанных процессов:
с одной стороны, увеличивается синхронность вовлечения в реакцию нервных клеток и длительность негативной фазы уменьшается, а с другой — одновременно происходит вовлечение в реакцию новых элементов, что способствует увеличению длительности потенциала. Но поскольку при локальной стимуляции пятнами небольших размеров микроочаг возбуждения в зрительной коре мозга крысы не превышает 200— 300 мкм [4], то и число вовлекаемых в реакцию нервных элементов в нем ограничено. Поэтому при увеличении размеров пятна более 7° длительность фокальных потенциалов в центре фокуса проекции уже не меняется. Описанные зависимости представлены на рис. 2.
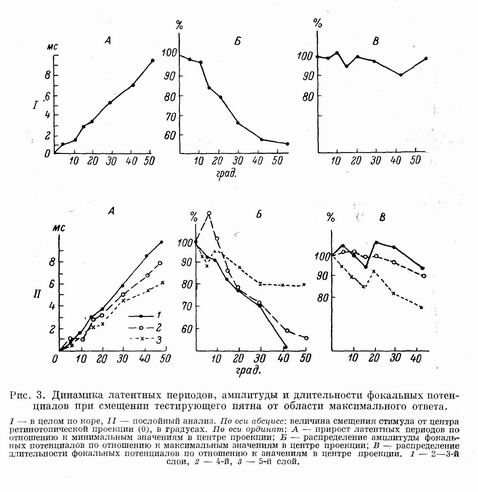
Распределение вызванных потенциалов при удалении стимулов от области максимального ответа.
При удалении тестирующего пятна от области максимального ответа увеличивается латентный период ответов. При смещении стимула на каждые 10: латентные периоды увеличивались в среднем на 1.8+0.15 мс. Амплитуда негативной фазы уменьшается. В среднем смещение стимулов на 10" от центра проекции приводит к падению амплитуды потенциала на 10+6%. Уменьшение длительности фокальных потенциалов было недостоверным.
Анализ динамики фокальных потенциалов по слоям коры мозга при удалении стимулов от области максимального ответа обнаружил более быстрое возрастание латентных периодов в верхних слоях в сравнении со средними и особенно нижними слоями. Изменение амплитуды и длительности при перемещении стимулов также зависит от слоя коры, в котором регистрируются ответы. Наиболее резко падает амплитуда потенциала в верхних слоях коры, но уменьшение длительности ответа было наименьшим. Обратная зависимость — малый декремент амплитуды и значительное укорочение длительности фокального потенциала наблюдается в 5-м слое.
Приведенные результаты находятся в соответствии с микроэлектродными исследованиями [2], в которых получены размеры возбужденных нейронных группировок при локальной световой стимуляции, составляющие около 200 мкм. Вот почему при удалении стимулов на 10° (примерно 200—250 мкм в коре мозга) изменения латентных периодов и амплитуды невелики (около 5%), а при перемещении стимулов на 20° от центра проекции эти изменения достигают уже 40%. Описанные зависимости приведены на рис. 3.
В заключение на основании полученных данных нами произведена пространственная реконструкция распределения возбуждения в поле 17 зрительной коры мозга при стимуляции локальными световыми пятнами (рис. 4).
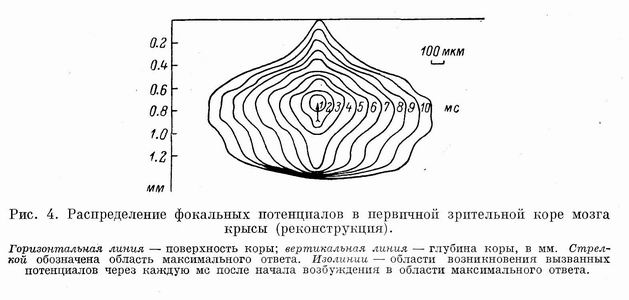
Таким образом, изучение распределения вызванных локальным светом потенциалов показало, что размеры фокуса возбуждения в воспринимающих слоях не превышают 200 мкм. Возбуждение соседних пунктов коры протекает при участии ряда последовательных и параллельных переключений.
ЛИТЕРАТУРА
[1] Артюхина Н. И. Влияние зрительной деафферентации на ультраструктуру синапсов зрительной коры крыс. Архив анат. гистолог, и эмбриол., 1975, 68, 4, 12-18.
[2] Ермаков П. Н. О функциональной организации нейронов зрительной коры мозга крысы при локальной и диффузной световой стимуляции, Автореф. дисс. Л. 1978.
[3] Коган А. Б. Вероятностно-статистический принцип нейронной организации систем мозга. ДАН СССР, 1964, 154, 1231—1235.
[4] Коган А. В., Ермаков П. Н. О формировании из микроочагов возбуждения мозаики активности в зрительной коре мозга крысы. Физпол. ж. СССР, 1977, 63, 5, 777—781.
15] Рокицкий П. Ф. Биологическая статистика. Минск, 1967.
[6] Супин А. Я. Нейрональные механизмы зрительного анализа. Изд. «Наука», М., 1974.
[7] Шевелев И. А. Динамика зрительного сенсорного сигнала. М., 1971.
[8] Adams A. D., Forrester J.M. The proection of rat's visual field on the cerebral cortex. Quart. J. Exper. Physiol., 1968, 53, 2, 327—336.
[9] Mont его V. M., Brugge J. F., Beitel R. E. Relation of the visual field to the lateral geniculate body of the albino rat. J. Neurophysiol., 1968, 31, 2, 221—237.
[10] Ruiz-Marcos A., Valverde F. Dinamic architecture of the visual cortex. Brain Res., 1970, 19, 25—39.
