/ Главная / Разные статьи / Радиационная биология
Статьи по радиационной биологии
Радиобиология”, т. XXV, вып.5, 1985, С.710-712.
К ВОПРОСУ О ВРЕМЕННЫХ ХАРАКТЕРИСТИКАХ ЛУЧЕВЫХ
РЕАКЦИЙ НЕЙРОНОВ СПИННОГО МОЗГА
ЩЕРБАТЫХ Ю. В.
Конечный эффект, регистрируемый в тканях после облучения, зависит от целого ряда факторов, одним из которых является время, прошедшее после облучения. Известно, что реализация лучевого поражения на клеточном уровне происходит поэтапно, проходя физическую, физико-химическую, химическую и другие стадии, которые, как правило, заканчиваются репродуктивной гибелью клеток [1]. В то же время для тканей, состоящих из малоделящихся или совсем не делящихся клеток (к таким системам относится сформировавшийся мозг), этот процесс может затягиваться на долгие месяцы и даже годы [2]. Это определяет ценность работ, в которых эффекты лучевого поражения рассматриваются в динамике, по мере их проявления. Как правило, такие исследования касаются нарушений мозгового кровообращения [3, 4] или поражения глии [5, 6], работы же, посвященные временным характеристикам лучевых реакций нейронной популяции мозга, носят единичный характер [7].
В опытах были использованы 24 крысы линии Вистар обоего пола массой 200— 250 г. Локальное облучение нижних поясничных сегментов спинного мозга в дозе 76 Гр проводили с помощью рентгеновского аппарата РУМ-17 (мощность дозы 0,12 Гр/с, напряжение 250 кВ, сила тока 15 мА, фокусное расстояние 21 см). Электрофизиологические и гистологические исследования проводили через 10, 25 и 50 суток после облучения. Использовали метод электрического раздражения периферического нерва парными стимулами с одновременной регистрацией потенциала дорзальной поверхности спинного мозга. После окончания электрофизиологического эксперимента облученный (или контрольный) участок мозга фиксировали в 12%-ном формалине, окрашивали гематоксилин-эозином, а затем подсчитывали измененные нейроны. Полученные результаты обрабатывали статистически с применением критерия Стьюдента
Исследования в области нейрорадиобиологии показали, что наиболее чувствительным к облучению звеном нейропиля является место специфического контакта нервных клеток — синапс [8]. Для изучения деятельности синаптического аппарата в физиологии применяется метод парных стимулов, подаваемых через короткие промежутки времени. Обычно синапсы способны в полной мере проводить возбуждение только при интервалах > 1 с, при меньших же интервалах проводящая способность синапсов падает, что выражается в снижении амплитуды постсинаптического потенциала. Этот процесс, описывающийся экспоненциальной кривой, является постоянным для данного вида синапсов и зависит от ряда факторов (количества выделяющегося медиатора, активности расщепляющих медиатор ферментов, чувствительности хемо- рецептивной мембраны и т. д.) [9].
При раздражении периферического, нерва n. tibialis сдвоенными электрическими стимулами нами было обнаружено, что после облучения вызванный потенциал на второй стимул был значительно меньше, чем у контрольных животных (рис. 1). Значительное уменьшение амплитуды второго ответа после облучения свидетельствует о том, что облучение ухудшает работу синапсов, причем степень этого ухудшения возрастает с течением времени, прошедшего после облучения. Более того, изменяется сам характер зависимости амплитуды потенциала от интервала между раздражениями: из экспоненциальной она переходит в линейную.
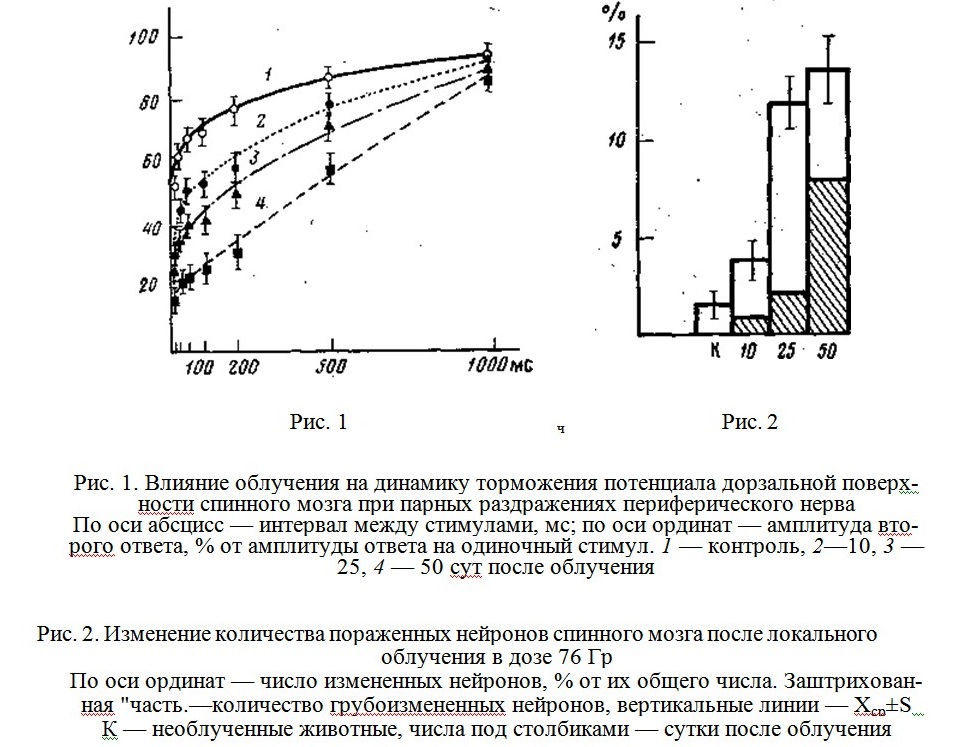
Чем чаще следовали друг за другом раздражающие импульсы тем заметнее были нарушения лабильности синаптических структурой только при очень редких раздражениях облученный синапс успевал восстанавливать свои функции и генерировать полноценный ответ на второй стимул.
Морфометрическое исследование препаратов облученного мозга выявило следующую картину (рис. 2), Через 10 сут после облучения число нервных клеток с признаками патологии составляло 3,7±0,9%, через 25 сут это число увеличивалось до 11,8±1,3%, причем преобладали слабоизмененные нейроны с сохранением нормального ядра, но со структурными изменениями цитоплазмы (набухание, гиперхроматоз, центральный хроматолиз и т. д.). Через 50 сут число поврежденных нейронов возрастало незначительно по сравнению с 25 сут (13,6±1,5%), зато степень поражения значительно увеличивалась и преобладающим становилось тяжелое изменение нейронов вплоть до полного распада клеток.
Полученные данные свидетельствуют о наличии тесной связи между развивающимися с определенным латентным периодом морфологическими и функциональными сдвигами в спинном мозге после локального рентгеновского облучения в дозе 76 Гр.
Литература
1.Кудряшов Ю. Б., Беренфельд Б. С. Основы радиационной биофизики. М.: Изд-во МГУ, 1982.
2.Baity О. Т., Woodard, J. S„ Putnam Т. J. Response of the nervous system to ionising radiation. Boston, 1964, p. 3—18.
3.Коgel A. J. van der. Radiation biology in cancer research. N. Y.: Raven Press, 1980, p. 61—470.
4.Reinhold H. S., Hopewell J.-W. Brit. J. Radiol., 1980, v. 53, p. 693—696.
5.Hubbard В. M., Hopewell I. W. Radiobiology, 1978, v 128, p. 779—781,
6.Zeman W. Proc. Nat. Acad. Sci., 1963, v. 50, p. 626—630.
7.Карповский A. JI., Федоренко Б. С., Рыжов H. И., Смирнова О. А. Радиобиология, 1981, т. XXI, вып. 3, с. 384—389,
8.Лебединский А. В., Нахильницкая 3. Н. Влияние ионизирующих излучений на нервную систему, М.: Атомиздат, 1960.
9.Eccles J. С., Rail W. J. Neurophysiol., 1951, v. 14, р. 353—376.
ON TIME-DEPENDENT RADIATION RESPONSES OF SPINAL CORD
SCHERBATYKH Yu. V.
State University named after the 300th Anniversary of Reuniting of the Ukraine and Russia, Dnepropetrovsk
In this report a description is given of functional and structural changes in spinal cord occurring 10, 25 and 50 days following local exposure to 76 Gy X-radiation. During the observation period, the capacity of synaptic structures was progressively impaired, the peripheral nerves irritated by high frequency, and neurons of exposed spinal cord segments changed.
**************
В. Е. Лебеденко, Ю. В. Щербатых, О. О. Шугуров. Изменения потенциалов спинного мозга крысы при тотальном облучении в дозе 360 гр. // Журнал «Радиационная биология. Радиоэкология», 1994, т.34. вып. 2. - С. 273-278.
V. E. Lebedenko, Yu. V. Shcherbatykh, О. О. Shugurov
CHANGES IN POTENTIALS OF SPINAL CORD UPON THE WHOLE-BODY IRRADIATION OF RATS
Research Institute of Biology, Dnepropetrovsk State University, Ukraine
In acute experiments in rats the effect of the whole-body X-irradiation with a dose of 360 Gy on induced somatosensory spinal cord potentials (SCP) was studied. The SSP components which show decrease in intesity and period depending upon a dose and time after irradiation were determined. The destruction of spinal input control systems was found. The causes and possible pathways of neuronal response depression which is probably due to synaptic structure damage have been considered. It is assumed that spinal potentials can be used as additive index of the extent of reaction to irradiation.
Аннотация:В острых опытах на крысах исследовали эффекты влияния' общего рентгеновского облучения в дозе 360 Гр на вызванные соматосенсорные потенциалы (ВССП) спинного мозга (СМ). Определены компоненты ВССП, амплитуда и длительность которых существенно уменьшаются в зависимости от дозы и времени после окончания облучения, показаны эффекты деструкции центральных систем регуляции на входе СМ. Рассмотрены причины и возможные механизмы угнетения нейрональных ответов, которое, вероятно, связано с поражением синаптических структур. Предполагается, что электрические потенциалы мозга могут служить дополнительным показателем величины лучевой реакции.
Ключевые слова: Спинной мозг, рентгеновское облучение, доза, нейроны, потенциалы, синапсы.
Известно, что вызванные соматосенсорные потенциалы (ВССП) спинного мозга (СМ), регистрируемые в его дорсальной части, отражают работу ряда нейронных систем на уровне сенсорных входов, моторных выходов и промежуточных интернейронов; осуществляющих взаимосвязь между входными и выходными цепями [1—3]. Электрическое развитие таких потенциалов обусловлено сомато-топической организацией нейронов СМ, включенных в возбуждаемые структуры [4,5].
В последнее время в медицинской и научной практике широко используются методы неинвазивной регистрации ВССП (с поверхности тела) для диагностики и анализа состояния СМ человека [2—4 ] или животных [6 ]. Поскольку абсолютно неприемлемо любое облучение человека, для моделирования указанных радиобиологических процессов в опытах были использованы крысы, так как их ВССП во многом схожи с ВССП человека [1, 6, 7]. Ранее было показано, что на фоне морфологических изменений нейронов некоторые компоненты таких потенциалов претерпевают значительные изменения вплоть до 50-х сут после локального облучения СМ в дозе 76 Гр [7 ]. Повышение дозы облучения до 360 Гр приводит к резкому сокращению сроков проявления морфологических изменений [8 ], которые, вероятно, находят свое отражение и в изменении электрофизиологических параметров данных потенциалов. Таким образом, целью данной работы явилось исследование характера и причин изменений электрофизиологических ^ реакций СМ в условиях тотального облучения в дозе 360 Гр.
МАТЕРИАЛЫ И МЕТОДИКА
Опыты были проведены на 19 крысах линии Вистар обоего пола массой 200—250 г. Общее облучение в дозе 360 Гр проводили с помощью рентгеновского аппарата РУМ-17 (мощность дозы 0,12 Гр/с, напряжение 250 кВ, сила тока 15 мА, фокусное расстояние 21 см). Электрофизиологические исследования спинного мозга (СМ) проводили через 1, 2, 3, 6, 7, 9 и 12 ч после облучения. Под нембуталовым наркозом (40—50 мг/кг внутрибрюшинно) проводили ламинэктомию в сегментах L6—S,, а также для раздражения препарировали нервы задней конечности (n. peroneus communis, n. tibialis communis), на которые наносили одиночные или парные раздражающие стимулы с силой 3 порога (относительно самых низкопороговых волокон). Вызванные соматосенсорные потенциалы (ВССП) СМ регистрировали по стандартной схеме с использованием методики накопления потенциалов на ЭВМ. Полученные результаты обрабатывали статистически с применением критерия Стьюдента.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Сразу же после облучения крысы находились в шоковом состоянии, были малоподвижны. Затем через 2—4 ч у них начинались судороги, развивалось двигательное беспокойство, которое заканчивалось параличом мышц задних конечностей. Все животные, облученные в дозе 360 Гр, погибали в течение 12 ч (в среднем через 8—9 ч).
Регистрация ВССП СМ, которая начиналась через 1—2 ч после облучения и велась вплоть до гибели животного, показала, что отдельные компоненты ВССП по-разному реагируют на применение сверхбольших доз радиации (рис. 1). Наиболее выраженным изменениям была подвержена позитивная (Р) волна, которая довольно быстро исчезала после облучения. Если у контрольных животных амплитуда Р-волны составляла в среднем 23±6% от наиболее высокоамплитудного первого негативного (N,) компонента ВССП, то спустя 2 ч она достигала только 6—7 %. Наряду с угнетением ее амплитуды отмечалось одновременное уменьшение и длительности — если в контроле она составляла 250—300 мс, то уже через 1 ч после облучения (рис. 1, К, 1) была менее 100 мс.
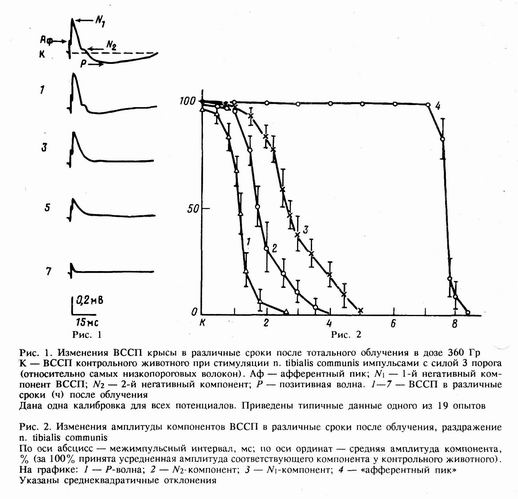
Спустя 2—4 ч после облучения практически полностью исчезал второй негативный компонент (N2), расположенный на нисходящей части N1- гo. Через 4—5 ч после облучения начинает угнетаться наиболее устойчивый, N1-компонент ВССП, что приводит к полному отсутствию постсинаптических компонентов. Следует отметить, что угнетение последних происходит без заметного сокращения их длительности.
Таким образом, через 7—8 ч после облучения при раздражении периферического нерва можно зарегистрировать только пресинаптический компонент ВССП — так называемый «афферентный пик», который исчезает последним (рис. 1,7). Это означает, что в разгар церебрального синдрома поступающая сенсорная информация полностью перестает обрабатываться в мозге. График изменений амплитуды основных компонентов ВССП в зависимости от времени после облучения представлен на рис. 2.
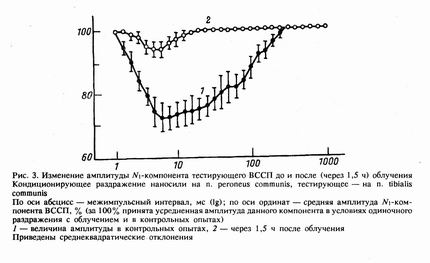
Согласно общепринятым моделям механизма пресинаптического торможения на входе СМ [9], исчезновение деполяризационного процесса в условиях стимуляции разных нервов двумя импульсами, разнесенными во времени, должно иметь следствием растормаживание второго по счету ответа. Это проверяли путем анализа величины ^-компонента ВССП. Изменение эффективности такого тррможения после облучения дозой 360 Гр можно заметить по величине амплитуды указанного компонента, вызванного тестирующим (идущим вторым в двойке) стимулом. При парных раздражениях двух различных периферических нервов (рис. 3) в интервалах от 5 до 250 мс наблюдали уменьшение угнетения тестирующего ответа.
В то время как одиночный ответ на раздражение п. tibialis communis через 1,5 ч после облучения составлял 95±3% от контрольного (т. е. не изменялся), торможение тестирующего ответа значительно уменьшалось. Если в норме (без облучения) величина тестирующего N1-компонента в максимуме торможения составляет 65—70% исходной, то через 1,5 ч после облучения — 90—95%. При увеличении времени облучения торможение отсутствовало почти полностью. Учитывая, что в случае раздражения двух разных периферических нервов гомонимные пути на тестирующий стимул практически не возбуждаются, можно говорить о том, что к 3-му часу пресинаптическое торможение полностью прекращает свою деятельность.
В своей оценке функциональных изменений ЦНС мы исходили из ранее полученных нами данных, а также данных других авторов о компонентном составе вызванных соматосенсорных потенциалов спинного мозга, а также о морфометрических исследованиях нейронов СМ при локальном и тотальном рентгеновском облучении [1,3, 6—8 ]. В настоящее время практически однозначно определены группы волокон, а также нейрональные механизмы, ответственные за генерацию известных компонентов ВССП. В частности, Р-волна связана с развитием пресинаптического торможения на входе СМ, N1-компонент генерируется возбуждением, пришедшим в мозг по афферентным волокнам группы А-бетта, N2-компонент — группы А гамма и дельта, «афферентный пик», являясь пресинаптическим компонентом, отражает возбуждение самых быстропроводящих низкопороговых волокон (группа A-альфа).
Как и предполагалось, увеличение дозы облучения с 76 до 360 Гр [7, 8 ] приводит к резкому увеличению скорости нарастания лучевой реакции в мозге. Хотя на клеточном уровне основные видимые изменения структуры приходятся на 3—6-й ч, функциональные изменения обнаруживаются уже через 1 ч после окончания воздействия. Это в первую очередь можно определить по угнетению всех полисинаптических реакций, ответственных за генерацию соматосенсорных потенциалов. Характерно, что, чем больше синаптических контактов в цепи генерации ответов, тем сильнее и быстрее угнетается сам ответ (рис. 2). Р-ъол ПДП, в генерации которой принимают участие нейроны желатинозной субстанции СМ (II—III слой по Рекседу) с обширными дивергентно-конвергентными связями, начинает деградировать уже в первые часы после облучения. Более крупные полисинаптические сегментарные нейроны, находящиеся в IV—VII слоях и ответственные за полисинаптические рефлекторные акты, угнетаются несколько позже — через 2—4 ч. Самыми устойчивыми из постсинаптических компонентов был высокоамплитудный N1-компонент, максимум депрессии которого приходился на 4—5 ч после облучения. Нейроны, генерирующие этот ответ, средней величины (15—20 мкм), находятся в IV—V слое СМ и моносинаптически связаны с волокнами группы А-бетта.
Можно предположить, что степень лучевой реакции (например, изменения ВССП) зависит от геометрических размеров нейронов СМ, поэтому первыми исчезают компоненты, связанные с маленькими клетками, — Р-волна, N2-компонент, а уж затем и N1. Однако это не так: проведенные исследования показали, что указанные компоненты имеют своеобразные параметры благодаря соматотопической организации нейрональных групп, генерирующих ответ, внутри объемного проводника — мозга [5 ]. Самые крупные нейроны СМ — мотонейроны, моносинаптически активируемые волокнами группы А-альфа, дающими «афферентный пик» ВССП, практически не вносят вклада в указанный потенциал (нет соответствующего компонента). Это прежде всего связано с недостаточной упорядоченностью электрических диполей, возникающих при возбуждении мотонейронов, и их сравнительно глубоком расположении в спинном мозге.
В связи со сказанным можно предположить, что наиболее чувствительной мишенью ионизирующего излучения является синапс — точка, где электрический сигнал преобразуется в химическую посылку. Аналогичное мнение было предложено и при анализе данных по локальному облучению мозга в дозе 76 Гр [7], а также в других исследованиях [10]. Это подтверждает наши данные о том, что самым устойчивым к облучению является пресинаптический компонент ВССП — «афферентный пик», отражающий приход потенциалов действия в СМ (т. е. до синаптического переключения). Следовательно, можно предположить, что мембранные процессы более устойчивы к внешнему физическому фактору, чем синаптические. Такое предположение может объяснить, почему резко уменьшаются амплитуда и длительность Р-волны ВССП, в то время как у моносинаптических ответов синхронность развития после облучения практически не изменяется, что выражается в постоянной длительности таких компонентов. Это подтверждают и данные о парных раздражениях разных периферических нервов с отведением N1-компонента (рис. 3). Поскольку при тестирующем раздражении активируются полностью другие (относительно кондиционирующего) афферентные волокна, в активацию вовлекаются и полностью новые синаптические контакты. Следовательно, растормаживание указанного ответа на фоне неизменного ^-компонента и «волоконного» компонента, каковым является афферентный пик ВССП, свидетельствует о синаптической природе этого феномена.
Таким образом, исходя из полученных результатов можно сделать следующие выводы.
1. На основании представленных и предыдущих [7, 8 ] опытов можно отметить, что увеличение дозы облучения сказывается на темпах угнетения электрических ответов мозга.
2. После общего облучения в дозе 360 Гр первоначально (через 1—3 ч) угнетаются электрические ответы мозга, связанные с возбуждением нейронов по полисинаптическим путям, несколько позже (4—5 ч) — по моносинаптическим.
3. Регуляторные механизмы обработки сенсорной информации, основанные на функционировании отрицательной обратной связи (например, пресинаптическое торможение в СМ), более чувствительны к облучению, чем механизмы прямых рефлекторных актов, что может определять динамику развития церебрального синдрома.
4. Изменения величины и длительности компонентов ВССП являются достаточно корректным показателем протекания лучевой реакции и могут использоваться для получения дополнительных данных о функционировании мозга после облучения.
СПИСОК ЛИТЕРАТУРЫ
1. Beall J. Е., Applebaum A. Е., Foreman R. D., Willlis W. D.lli. Neurophysiol. 1977. V. 40. № 2.
P. 199—211.
2. Cole J. D., Katifi H. A., Sedgwick E. M.lli. Physiol. (Gr. Brit.). 1985. V. 358. P. 34.
3. Cracco R. 0.//Electroenceph. and Clin. Neurophysiol. 1973. V. 35. P. 379—386.
4. Delbeke J., Me Comas A. J., Kopec S. J.III. Neurol. Neurosurg. Psychiatr. 1978. V. 41. P. 293—302.
5. Brown A. C.//J. Exp. Physio!. 1982. V. 67. № 1. P. 193—212.
6. Шугуров О. О. Биологические и технические системы управления. Днепропетровск: Изд-во ДГУ,
1984. С. 18—21.
7. Шербатых Ю. В.//Радиобиология. 1985. Т. 25. Вып. 5. С. 710—712.
8. Щербатых Ю. В., Клеринг П. Г., Дворецкий А. //.//Радиобиология. 1986. Т. 26. Вып. 3. С. 397—399.
9. Экклс Дж. Физиология синапсов. М.: Мир, 1966. 396 с.
10. Ткаченко 3. Я., Демидова Г. Г.//Радиобиология. 1974. Т. 14. Вып. 4. С. 621—624.
Влияние исходного функционального состояния на лучевую реакцию нейронов спинного мозга крысы. О.О. Шугуров, Ю.В. Щербатых. Радиационная биология. Радиоэкология. 1998, т.38, вып.3, С. 389-392.
О.О. Shugurov, Yu.V. Shcherbatykh. Relation between basis functional state of spinal cord and morphometrical parameters of neurons upon local x-radiation.
Scientific Research Institute of Biology of Dnepropetrovsk Stale University, Ukraine
The influence of CNS functional state and structural changes of neurons in spinal cord following local exposure to 38 or 76 Gy X-radiation. Morphological analysis show, that stimulation of peripheral nerves increase, but hypoxia or barbiturates decrease destruction of spinal neurons by radiation. Value destruction also depends of neurons volumes.
Аннотация:Исследовали влияние функционального состояния ЦНС на степень ее поражения рентгеновским излучением в дозах 38 и 76 Гр. В качестве факторов, изменяющих состояния ЦНС, использовали наркоз, гипоксию, электрическую стимуляцию периферических нервов, проводили морфометрические измерения параметров нейронов спинного мозга. Анализ полученных данных показал, что повышенная активность ЦНС в период облучения ведет к увеличению патологических изменений нейронов (в первую очередь мелких и средних), угнетение нейрональной активности в период облучения (наркоз, гипоксия) проявляет протекторные свойства. Сделано предположение о возможности использования тормозных медиаторов в качестве специфических радиопротекторов ЦНС.
Ключевые слова: Нейрон, спинной мозг, облучение, морфометрия, радиопротектор.
Одной из причин затрудняющих сопоставление результатов клинических и лабораторных экспериментов, является то, что лучевая терапия опухолей нервной системы осуществляется у бодрствующих людей [1], в то время как локальное облучение животных, производимое для отработки или проверки новых методов лечения, в силу ряда методических особенностей происходит при их иммобилизации барбитуратами [2, 3], а при фракционированном облучении - ингаляционными анестетиками [4, 5]. Ранее была высказана мысль, что барбитураты угнетают физиологическую активность нервных клеток, повышая в связи с этим их радиорезистентность [6]. Активное внедрение в радиологическую практику газовой гипоксии [10, 11] требует детального изучения этого воздействия на нервную систему. Таким образом, в задачу исследования входило изучение влияния исходного состояния нервной ткани на изменения, вызванные локальным рентгеновским облучением.
МАТЕРИАЛЫ И МЕТОДИКА
Опыты проводили на 23 крысах линии Вистар обоего пола массой 200—250 г. Локальное облучение в дозе 38 или 76 Гр проводили с помощью аппарата РУМ-17 (мощность дозы 0,12 Гр/с, напряжение 250 кВ; сила тока 15 мА, фокусное расстояние 21 см) с использованием описанного ранее устройства для локализации воздействия [7].
Все исследования проводили на 30-е сутки после облучения. Для оценки степени лучевой реакции использовали морфометрические параметры нейронов до и после облучения в различных условиях функционального состояния (животное под наркозом, в условиях гипоксии, электрической стимуляции соматосенсорных зон под наркозом, контроль). Наркоз применяли в дозе 75 мг/кг веса, гипоксию определяли газовым составом воздушной смеси (8% О2 и 92% N2), стимуляцию под указанным выше наркозом осуществляли на п. tibialis посредством предъявления ритмической серии импульсов длительностью 0,3 мс и частотой 20 1/с (в течение облучения). Контроль — необлученное животное, норма - облученное животное в установке по локальному облучению [7] без использования исследуемых нагрузок. Измерение параметров нейронов осуществляли на аппарате «Мофоквант», их анализ проводили на ЭВМ с применением критериев достоверности.
Морфологические показатели нейронов спинного мозга через 1 мес. после облучения в дозе 38 Гр.
|
Группа животных
|
Отношение малой и
большой осей нейронального
ядра
|
Отношение объема
цитоплазмы к поверхности ядра (мкм3/мкм2)
|
|
|
|
|
1
|
|
2
|
1
|
|
2
|
|
Необлученные
животные
|
0,78±0,02
|
0,78±0,02
|
25,9±1,5
|
12,7±1,0
|
|
Облученные животные
|
0,74±0,02
|
0,70±0,02
|
36,1±2,7
|
16,7±2,6
|
|
Облученные +гипоксия
|
0,76±0,04
|
0,77±0,03
|
26,4±3,0
|
14,4±2,8
|
|
Облученные +наркоз
|
0,77±0,5
|
0,77±0,08
|
26,3±2,2
|
13,8±1,9
|
|
Облученные +стимуляция
|
0,73±0,3
|
0,68±0,2
|
38,2±3,0
|
16,9±3,3
|
Примечание. 1 - крупные нейроны с длинной осью более 35 мкм, 2 - средние нейроны с длинной осью до 35 мкм.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Морфометрические исследования показали, что общее число измененных нейронов вентральных рогов спинного мозга (СМ) крыс, облученных в бодрствующем состоянии, составляло 4,5±1,1% при дозе 38 Гр и 12,3+1,3% при дозе 76 Гр. У облученных под наркозом — 5,2+1,4 и 1,8+1,2% соответственно. Изучение гистологических образцов нервной ткани крыс, забитых через 1 мес после локального облучения в условиях газовой гипоксии, показало 3,6+1,2% измененных клеток при дозе 38 Гр и 10,5+1,6% при дозе 76 Гр. В то же время общее число измененных клеток СМ при воздействии облучения на фоне электростимуляции распределяется как 6,2±2,6 и соответственно 16,7+3,1%.
Следует отметить, что увеличение дозы облучения увеличивало абсолютный разброс в данных, тем не менее общая картина изменений становилась более отчетливой. При этом суммарные изменения нейронов оценивались по целому ряду параметров: внутриклеточному смещению ядра, площади сечения нейронов, объему клетки, площади поверхности и объему ядра, коэффициенту сжатия ядра и ядерно-цитоплазмати-ческому отношению. В таблице даны примеры изменения величин двух морфометри-ческих параметров в условиях облучения в дозе 38 Гр. Суммарные же изменения, выраженные в процентах для двух доз, представлены на рисунке.
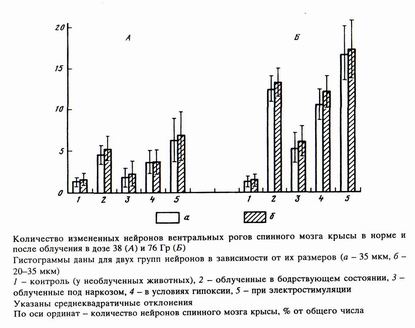
Наибольшие изменения морфометрических величин наблюдаются на средних и мелких нейронах спинного мозга и менее значительные у крупных нейронов (в основном мотонейронов). Учитывая данные по необлученным нейронам, можно считать, что количество достоверно измененных нейронов в случае облучения на фоне прямой стимуляции нервов максимально и достигает 20% их общего количества. У облученных животных без изменения условий (норма) деградация формы и параметров отмечена у 15% нейронов. В то же время гипоксия и тем более наркоз приводят к относительному ослаблению изменений морфометрических параметров при последующем рентгеновском облучении. Кроме того, заметно, что увеличение дозы облучения в 2 раза ведет к почти троекратному увеличению негативных последствий, что свидетельствует о нелинейном характере зависимостей доза — морфометрическое изменение параметра нейрона.
Результаты опытов подтверждают данные литературы, согласно которым барбитураты уменьшают степень лучевого поражения нервной системы [8, 9], хотя механизм этого явления полностью не раскрыт. Мнение некоторых авторов, что нембута-ловая анестезия может изменять радиочувствительность за счет создания тканевой гипоксии [5], вряд ли является исчерпывающим в отношении нервной ткани, так как более выраженный протекторный эффект наркоза по сравнению с гипоксией, отмеченный как в наших опытах, так и литературе [6], свидетельствует о специфическом характере взаимодействия барбитуратов с нервной тканью во время облучения.
Наши предыдущие данные свидетельствуют о том, что морфологическим изменениям сопутствуют изменения функциональные, которые находят свое отражение в ряде массовых потенциалов спинного мозга [12, 13]. Опыты показывают, что активация нервных структур "под лучом" приводит к усилению нейротрансмиттерных реакций, что ведет к существенным негативным морфофункциональным изменениям. И наоборот, затормаживание активности с помощью барбитуратов или гипоксии уменьшает вероятность протекания биохимических реакций, тем самым сокращая диапазон мишеней рентгеновского облучения. Но именно такие функции могут выполнять нейрогормоны и нейромедиаторы, осуществляющие в спинном мозге активное торможение биоэлектрических реакций. Используя такой подход, можно предположить, что применение таких тормозных медиаторов, как ГАМК или глицин, перед облучением СМ может существенно снизить негативность облучения ЦНС.
СПИСОК ЛИТЕРАТУРЫ
1. Бадмаев К.И., Смирнов F.B: Радионуклеидная диагностика и лучевая терапия заболеваний нервной системы. М.: Медицина. 1982. 132 с.
2. Zeman W. // Proc. Nat. Acad. Sci. 1963. V. 50. P. 626-630.
3. Knowles J.S. II Int. J. Radiat. Biol. 1983. V. 44. № 5. P. 433-442.
4. Reinhold H.S., Hopewell J.W. II Brit. J. Radiol. 1980. V. 53. P. 693-696.
5. AngK.. KogelA., Schuger E. //Int. J. Rad. Oncol. Biol. Phys. 1983. V. 9. № 10. P. 1487-1491.
6. Минаев П.Ф. Влияние ионизирующего излучения на ЦНС. М.: Изд-во АН СССР, 1962. 200 с.
7. Щербатых Ю.В., Шиш Д.И. Биологические и технические системы управления. Днепропетровск: Изд-во ДГУ, 1984. С. 37-38.
8. AlwordE.C, Brace КС. Hi. Neuropathol. 1957. V. 16. № 1. P. 3-17.
9. Семенов Л.Ф. II Мед. радиология. 1958. Т. 3. № 3. С. 70-77.
10. Чижов А.Я. Механизмы противолучевого действия гипоксической гипоксии и экспериментально-клиническое обоснование ее использования для ослабления лучевого поражения при общем и локальном облучении организма: Автореф. дис. ... д-ра мед. наук. Обнинск, 1983. 32 с.
11. Ярмоненко СП. Радиобиология человека и животных. М.: Высш. шк., 1984. 376 с.
12. Щербатых Ю.В. // Радиобиология. 1985. Т. 25. Вып. 5. С. 710-712.
13. Лебеденко B.E., Щербатых Ю.В., Шугуров О.О. Радиац. биология. Радиоэкология. 1994. Т. 34. Вып. 2. С. 273-277.
